A Conceptual Introduction:
The study of genetics reveals a stunning tapestry of diversity within the living world. While evolutionary theory traditionally attributes this variation to random mutations accumulated over vast stretches of time, a creationist perspective offers a compelling alternative: Created Heterozygosity. This hypothesis proposes that God designed organisms with pre-existing genetic variability, allowing for adaptation and diversification within created kinds. This concept not only aligns with biblical accounts but also provides a more coherent explanation for observed genetic phenomena.
The evolutionary narrative hinges on the power of mutations to generate novel genetic information. However, the overwhelming evidence points to the deleterious nature of most mutations. This can be seen in the famous Long-Term Evolutionary Experiments with E. coli. Notice, in the graphic below (Hofwegen, 2016), just how much information gets lost due to selection pressures and mutation. This is known as genetic entropy, the gradual degradation of the genome due to accumulated harmful mutations, poses a significant challenge to the idea that random mutations can drive the complexification of life. Furthermore, the sheer number of beneficial mutations required to explain the intricate design of living organisms strains credulity.
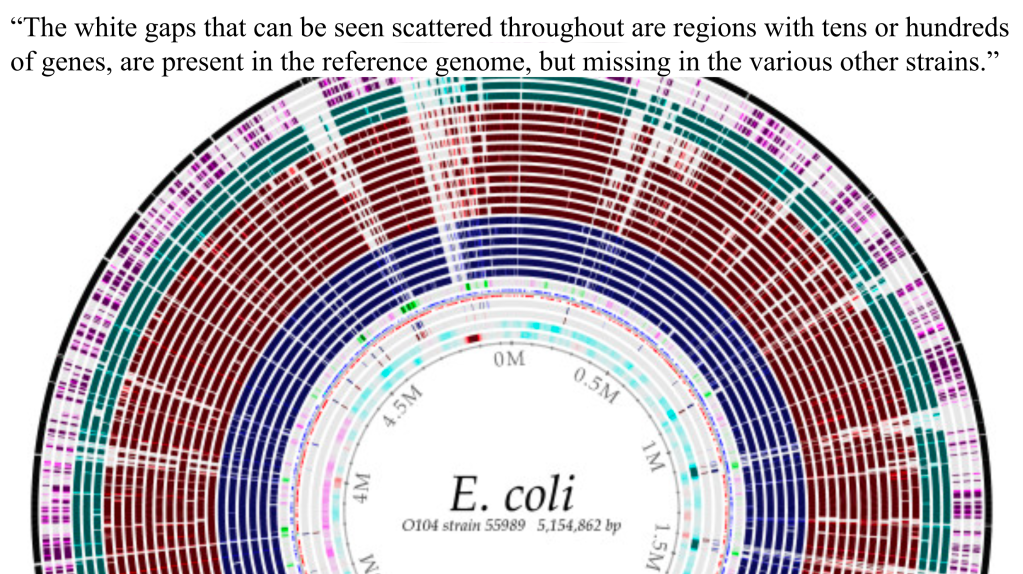
“Genomic DNA sequencing revealed an amplification of the citT and dctA loci and DNA rearrangements to capture a promoter to express CitT, aerobically. These are members of the same class of mutations identified by the LTEE. We conclude that the rarity of the LTEE mutant was an artifact of the experimental conditions and not a unique evolutionary event. No new genetic information (novel gene function) evolved.”
In contrast, Created Heterozygosity suggests that God, the master engineer, imbued organisms with a pre-programmed potential for variation. Just as human engineers design systems with built-in flexibility, God equipped his creation with the genetic resources necessary to adapt to diverse environments. This concept resonates with the biblical affirmation that God created organisms “according to their kinds,” implying inherent boundaries within which variation can occur. Recent research, such as the ENCODE project and studies on the dark proteome, has revealed an astonishing level of complexity and functionality within the genome, further supporting the idea of a designed system.
Baraminology, the study of created kinds, provides empirical support for Created Heterozygosity. The rapid diversification observed within baramins, such as the canid or feline kinds, can be readily explained by the expression of pre-existing genetic information. For example, the diverse array of dog breeds can be traced back to the inherent genetic variability within the canine kind, rather than the accumulation of countless beneficial mutations.
Of course, objections arise. The role of mutations in adaptation is often cited as evidence against Created Heterozygosity. However, certain mutations may represent the expression of designed backup systems or pre-programmed responses to environmental changes. Moreover, the vast majority of observed genetic variation can be attributed to the shuffling and expression of existing genetic information, rather than the creation of entirely new information.
The implications for human genetics are profound. Created Heterozygosity elegantly explains the high degree of genetic variation within the human population, while remaining consistent with the biblical account of Adam and Eve as the progenitors of all humanity. Research on Mitochondrial Eve and Y-Chromosome Adam/Noah further supports the idea of a recent, common ancestry for all people.
In conclusion, Created Heterozygosity provides a compelling framework for understanding genetic variation from a creationist perspective. By acknowledging the limitations of mutation-driven evolution and recognizing the evidence for designed diversity, we can appreciate the intricate wisdom of the Creator and the coherence of the biblical narrative. This concept invites us to explore the vastness of genetic diversity with a renewed sense of awe, recognizing the pre-programmed potential inherent in God’s magnificent creation.
Citation:
- Van Hofwegen, D. J., Hovde, C. J., & Minnich, S. A. (2016). Rapid Evolution of Citrate Utilization by Escherichia coli by Direct Selection Requires citT and dctA. Journal of bacteriology, 198(7), 1022–1034.